We, as musicians and/or fans of music, rarely engage with the highly tuned mechanism lodged within our skull – the ear. We take full advantage of our ability to use our ear not only as a survival tool but also a medium for entertainment, but we rarely ever think about how it works. This particular blog will provide a quick guide through Auditory Transduction – how we perceive acoustical energy, how we transform it into electromechanical energy, and how we process that information biologically. Once we have a better understanding of that process we will also revisit the Absolute Pitch discussion and illustrate some new material you will find interesting.
Perception – What do you hear?
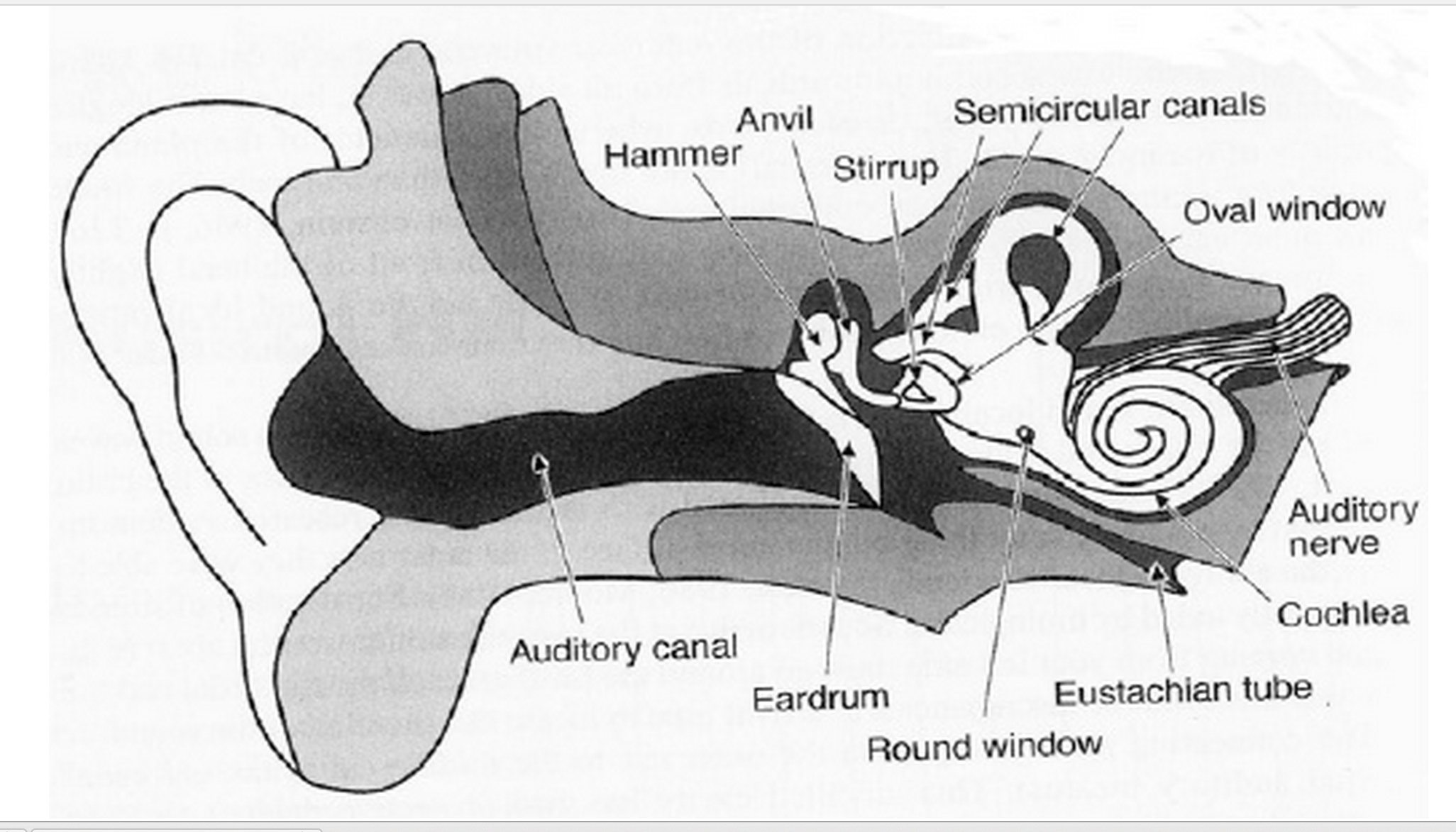
The depiction below illustrates the construction of the outer, middle, and inner ear (Hodges & Sebald 2011, p. 98).
Brandon Pletch provides a wonderful video describing the Auditory Transduction process. I will provide as shortened description here but it is best for you to watch the video and pick back up in the next section.
Outer Ear (air filled)
Acoustical energy (sound) first enters the ‘outer-ear’ by way of the pinna (ear lobe) and travels through the external auditory meatus (ear canal) striking the tympanic membrane (ear drum).
Middle Ear (air filled)
When acoustical energy strikes the tympanic membrane it is converted to mechanical energy through the use of three small bones. The malleus (hammer) – attached to the tympanic membrane, the incus (anvil), and the stapes (stirrup) – attached to the oval window.
Inner Ear (fluid filled)
The oval window separates the air-filled middle ear from the fluid-filled inner ear. The inner ear consists of the semicircular canals and the cochlea. The semicircular canals act as our gyroscope, determining body position. They are not involved in the process of hearing.
The cochlea is divided into two parts: the scala vestibuli (vestibular canal) and the scala tympani (tympanic canal) which are connected at the tip by the helicotrema. The membrane separating the canals is called the cochlear duct. This duct contains the Reissner’s membrane and the Basilar membrane. The organ of Corti, attached to the Basilar membrane, is where the mechanical energy that enters the oval window is converted tonotopically (frequency dependent) into electromechanical energy through the use of the hair fibers which send the newly converted neural signal to the auditory nerve.
Ear to Brain
The brain is immensely complicated. My intent is to give you a very basic idea of how musical information from the ear is ingested and processed by the brain. Hodges and Sebald (2011, p. 102) provides a general overview of the transmission process:
(1) Analysis of auditory information takes place in many localized processing centers scattered along a diffuse pathway between the inner ear and the auditory complex.
(2) Each of these processing centers analyzes the auditory information for a particular feature – location, source etc…
(3) All of these bits of information are put back together to create a coherent sound experience with meaning in the primary auditory cortex and surrounding associating areas (below in yellow).

The nerve impulse from the ear travels to the brain stem through a series of processing structures and arrives at the Auditory Cortex. I would like to draw your attention to a specific part of the auditory cortex named the Heschel’s Gyrus. Dr. Charles Limb in his study, Structural and Functional Neural Correlates of Music Perception asserts that while one might expect higher levels of neural processing to be most intrinsically related to musical processing, Heschel’s gyrus itself has received a good deal of attention as a possible marker of musicality.
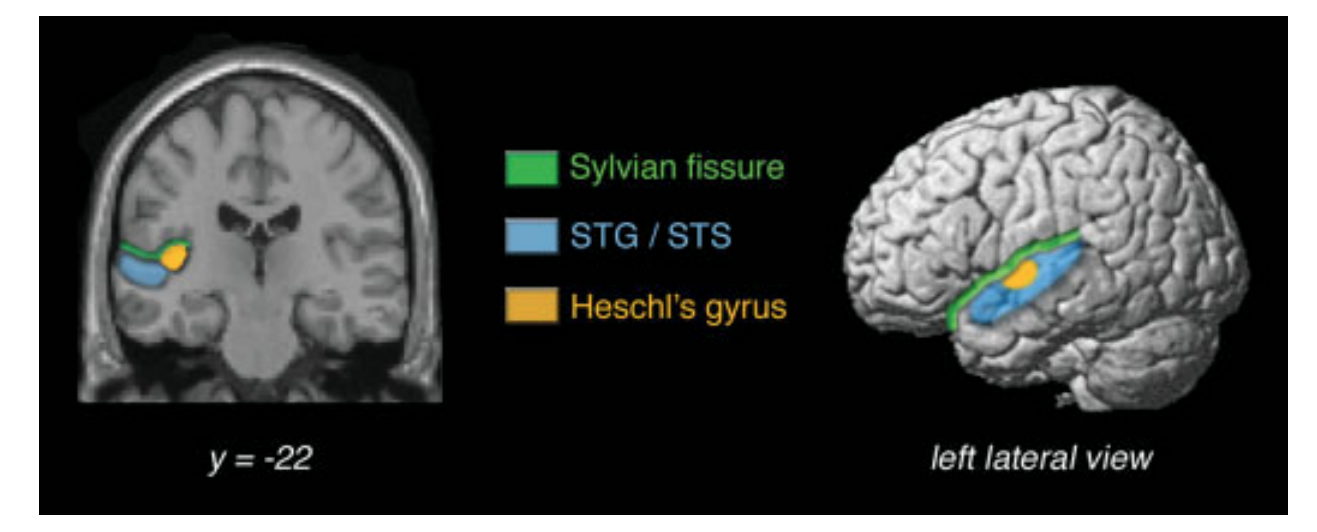 Limb’s study found that professional musicians showed a significant (greater than 100%) increase in MEG activity within primary auditory cortex compared to nonmusicians, which was found to correlate with increased (130%) volumetric measurements of gray matter within Heschl’s gyrus in musicians compared to nonmusicians. Furthermore, psychometric testing revealed a positive correlation between the size of Heschl’s gyrus and musical aptitude. (p. 437).
Limb’s study found that professional musicians showed a significant (greater than 100%) increase in MEG activity within primary auditory cortex compared to nonmusicians, which was found to correlate with increased (130%) volumetric measurements of gray matter within Heschl’s gyrus in musicians compared to nonmusicians. Furthermore, psychometric testing revealed a positive correlation between the size of Heschl’s gyrus and musical aptitude. (p. 437).
Further discussion concerning the brain can be found towards the end of this blog post.
Absolute Pitch Revisited
As discussed in the first blog post concerning this topic matter, Absolute Pitch (AP) is the ability to identify the pitch of tones without the use of any external reference. Cognitive theorists suggest that even if genetics play a role, existing research suggests that a critical learning period is involved. This “research” is alluding to assertions that one of the best generalizations on can make about “perfect pitch” (AP) is that its possessors typically begin musical instruction or involvement at a comparatively early age – often before the age of six or seven years. Assertions like this are becoming less popular. Here we will review Pitch Memory and Pitch Labeling, then go into material prepared by Dr. Ian Quinn including the Two-Mechanism Theory by Dr. David Ross, and also get back to the brain and discuss newer research findings supporting Quinn’s lecture.
Pitch Memory and Pitch Labeling
Daniel Levitin put together an article regarding a study concerning learned melodies. Within the article, he explained what is called the Two-Component Theory of Absolute Pitch. Levitin postulates that AP is merely a small extension of memory abilities that are widespread in the music population and AP consists of two distinct component abilities: (1) Pitch Memory and (2) Pitch Labeling.
Pitch Memory is the ability to maintain stable, long-term representations of specific pitches in memory and to access them when required (think Computer RAM). Pitch Labeling is the ability to attach meaningful labels to pitches. This attachment is thought to be coded implicitly through an alternate set of perceptual categories (perhaps the genetic advantage?) used in sound processing in Long Term Memory. Below is a diagram put together by Cognitive Theorist, Dr. Ian Quinn while lecturing at the University of Colorado in November of 2014. The diagram outlines (in simple illustrations) where the populace lies in regards to having non-absolute pitch (NAP) or absolute pitch (AP). For one to truly have absolute pitch it would be appropriate, if not mandatory, that an AP user possess the cognitive proclivity for both pitch memory and pitch labeling.
(Click on the picture for better resolution)

It is important to know that if you do possess AP do not simply rely on it to help you with intervallic relationships and function. To put it another way, identifying pitch out of context is irrelevant and even meaningless to music (Miyazaki 1993).
Two Mechanism Theory of Absolute Pitch (Ross 2005)
In a study titled, Absolute Pitch Does Not Depend on Early Musical Training, Drs. David Ross, Ingrid Olson, and John Gore concludes that musical training is not necessary for the development of AP.
Two Mechanism Theory (Ross 2005):
(1) Ability to Perceptually Encode Pitch
– A basic property of perceptual encoding
– Durable representations of stimulus frequency are formed at a pre-categorical level
– Likely innate
– Does not come in degrees
(2) Heightened Tonal memory
– A skill of retrieval and comparison
– Increased ability to retrieve specific complex auditory memories into working memory
– Likely learned
– Comes in degrees
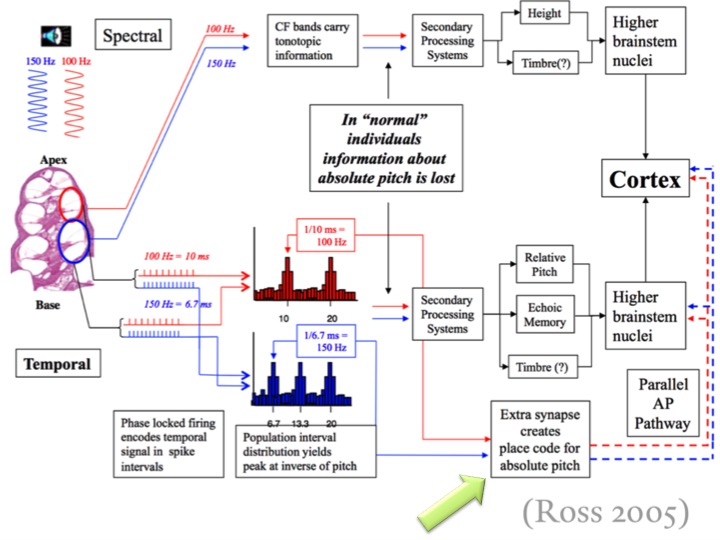
In the chart above (provided by Dr. Quinn), Ross spells out the acoustical signal pathway to the Auditory Cortex. Pay special attention to the green arrow. Ross and Quinn both indicate that there is a special neural pathway that is specifically used to encode (chunking, schemata), rehearse, and store data having to do with AP. It is this special neural network within which the brain retrieves information in regards to instances where AP dependency is called upon. This information strongly suggests that AP is a genetically determined trait. Ross further concludes that both musicians and non-musicians (unable to name note names) with AP faired far better than their non-AP possessors throughout the study disproving the hypothesis that early musical training is required for the development of AP.
Here is where things get fun! To this point I have argued for the position that AP is a genetically determined. I am also going to take the position that early musical training helps develop (not create) this genetic trait.
Planum Temporale and Pruning
Earlier, we spoke about the brain on a basic level including the Heschel gyrus. I also want to bring the Planum Temporale (PT) to your attention. It is a portion of the brain seated next to the anterior temporal lobe of the brain.
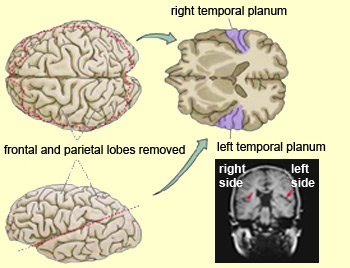
This region is associated with language and auditory processing. The results of a study conducted by Drs. Keenan, Thangaraj, Halpern, and Schlaug titled, Absolute Pitch and the Planum Temporale (2001) sought to better understand why the PT is asymmetrical with the left PT larger than the right in those who have AP found in earlier studies with smaller samples – Zatorre (1989) and Schlaug et al. (1995). Keenan et al. found that not only was the left hemisphere PT larger in AP possessors but the right hemisphere PT was dramatically smaller (below). Most humans inherently have a larger left hemisphere PT. It is the right PT that is of most interest.

Keenan et al. conclude that their data suggest that early developmental pruning in the right PT may create an anatomical dominance of the left PT. This, in turn, might create a functional dominance of the left PT over the right PT, which might be necessary for the acquisition and/or manifestation of absolute pitch. Keenan’s summary takes a confusing turn when he mentions the following, “This anatomical difference is likely due to factors other than early musical training or music exposure and it might indicate that young children with an increased leftward PT asymmetry might develop AP if they have an early music exposure.” This sentence aside, it is apparent from the study and imaging that AP is correlated with the decreased size of the right PT.
Pruning
Throughout the first four to eight years of life, the brain will have produced far more synapses than is necessary for a human’s lifetime. The reason for the overproduction of synapses is that biologically the brain is preparing to meet any environmental challenge it might encounter (Hodges & Sebald 2011). Genetic instructions and learning experiences work together to sculpt a child’s brain into its eventual adult configuration. It is at this point where the determination of the synaptic structure of the PT’s are determined lending credibility to the genetic proclivity towards possessing AP.
From the findings I have gathered, AP can not be developed late in life. It is a trait resulting in an alternate set of neural pathways developed and pruned at an early age without the necessity of musical training, but if one happens to possess AP, they are far more likely to develop that trait through the process of early musical training. As I have stated in previous blogs, reliance on that trait is unnecessary and could also be a hindrance when making music with others who do not possess AP – which turns out to be 9,999 out of 10,000 people out there.
Jason Michael Johnston
DMA Candidate, University of Colorado at Boulder
#muscog
References
Hodges, Donald A. and David C. Sebald. Music in the Human Experience: An Introduction to Music Psychology. New York: Routledge, 2011.
Keenan, Julian P.,Van Thangaraj, Andrea R. Halpern, Gottfried Schlaug. “Absolute Pitch and Planum Temporale.” NeuroImage, 2001, 14, 1402-1408.
Levitin, Daniel. “Absolute Memory for Musical Pitch: Evidence from the Production of Learned Melodies.” Perception & Psychophysics, 1994, 56 (4), 414-423.
Limb, Charles J. “Structural and Functional Neural Correlates of Music Perception.” The Anatomical Record, 2006, 288A (4), 435-446.
Miyazaki, Ken’ichi. “Absolute Pitch as an Inability: Identification of Musical Intervals in a Tonal Context.” Music Perception: An Interdisciplinary Journal, 1993, Vol. 11, No. 1, 55-71.
Ross, David A., Ingrid R. Olson, and John C. Gore. “Absolute Pitch Does Not Depend on Early Musical Training.” Annals of the New York Academy of Sciences, 2003, 999, 522-526.
